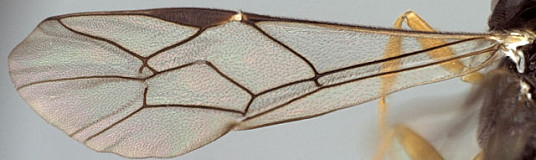
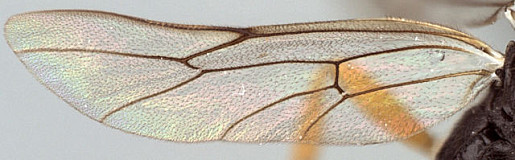
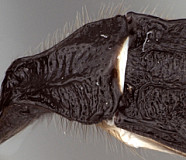
Antipygus Tschek, 1868: 438. Type species: Antipygus megerlei Tschek. Monobasic. Synonymized by Townes et al. (1965: 241).
Holmgren (1855) is sometimes listed under various dates.
Antipygus Tschek, 1868: 438. Type species: Antipygus megerlei Tschek. Monobasic. Synonymized by Townes et al. (1965: 241).
Holmgren (1855) is sometimes listed under various dates.
The above description is modified from Townes (1970) and was based largely on three North American species in the Texas A&M University Collection.
Acantholyda erythrocephala Pinus (Aubert 2000)
Acantholyda hieroglyphica (Ozols 1961)
Acantholyda posticalis (Hardtl 1943; Aubert 2000)
Acantholyda posticalis Pinus (Shaw et al. 2003)
Cephalcia (Martinek 1989)
Cephalcia abietis (Sedivy 1965, Sedivy 1967; Jahn 1978; Eichhorn 1988; Martinek 1989; Martinek 1990; Kanecka 1993, 1995; Eichhorn and Bogenschutz 2000; Kasparyan 2002)
Cephalcia alpina (Rohrig 1953)
Cephalcia arvensis (Boas 1933; Martinek 1991, 1992)
Cephalcia arvensis fallenii (Martinek 1992)
Cephalcia lariciphila (Aubert 2000)
Pamphilius depressus (Hinz 1961)
Pamphilius hortorum (Hinz 1961)
Pamphilius hypotrophicus (Strand 1915; Scheidter 1916; Hedwig 1962; Schimitschek 1964)
Pamphilius latifrons (Shaw et al. 2003)
Pamphilius pallipes ( Kasparyan 2002)
Pamphilius pallipes Betula (Shaw et al. 2003)
Pamphilius vafer (Villemant 1982; Aubert 2000; Kasparyan 2002)
The following host records are doubtful.
Cimbex (Rondani 1872)
Cimbex femorata (Starke 1956)
Cydia strobilella (Constantineanu and Istrate 1971; Pisica and Petcu 1972)
Tenthredo campestris (Ratzeburg 1844)
Thaumetopoea pityocampa (Romanyk and Ruperez 1960)
Trichiosoma betuleti (Rudow 1917Hedwig 1958)
Trichiosoma nanae (Vikberg and Viitasaari 1991)
Trichiosoma sachalinense (Aubert 2000)
Trichiosoma sorbi (Rudow 1882)
There are no specimens currently determined for this OTU, or those specimens determined for this OTU are not yet mappable.
The keys, text, and images presented here were initially developed as part of an NSF-sponsored PEET project, DEB0328922. This page is a joint project of NSF PEET grants NSF DEB-0328922; ideas and opinions expressed here are those of the authors, and not the NSF; best viewed with Firefox, Mozilla, or OS X Safari. This material is copyright Bob Wharton (2007 and onwards) and may not be reproduced without permission.