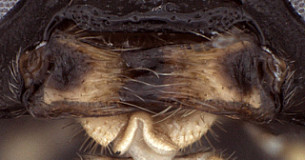
Clypeus with surface more or less uniformly setose and finely rugulose (Fig. 2); ventral margin evenly but weakly convex, the margin blunt but not broadly so (Fig. 2); epistomal sulcus impressed throughout, weaker medially; clypeus in profile weakly protruding (Fig. 1). Face setose, densely granular-punctate medially, faintly punctate, nearly smooth laterally as well as on frons and vertex; inner eye margins weakly (Fig. 2) to distinctly converging. Malar space short but distinct, about half basal width of mandible in female, slightly longer in male; malar sulcus absent. Mandible (Fig. 3) broad, nearly parallel-sided; dorsal tooth distinctly narrower and a little shorter than ventral tooth; basal, transverse impression absent; ventral margin distinctly carinate. Maxillary palp a little shorter than height of head; female antenna slightly shorter than body; about equal in length to body in male. Hypostomal carina meeting occipital carina at base of mandible or the two very slightly separated at the base; occipital carina complete. Epicnemial carina reaching anterior margin of mesopleuron. Notaulus distinct, varying from weakly to deeply impressed, sculptured when deeply impressed; extending posteriorly to level of tegula. Groove between propodeum and metapleuron broadly u-shaped, readily visible in lateral view; broad, u-shaped groove mid-dorsally between propodeum and metanotum also readily visible in lateral view; pleural carina well-developed, complete; median and lateral longitudinal carinae well developed anteriorly, variously effaced posteriorly, though rarely completely so; posterior transverse carina sometimes complete laterally, usually absent medially, anterior transverse carina present medially absent laterally, thus areola usually open posteriorly. Hind femur stout; apical margin of mid tibia not expanded into a distinct tooth similar to that of fore leg; apical comb on posterior side of hind tibia absent; posterior hind tibial spur about 7x longer than maximum width at base; tarsal claws smaller than in
Pion, not pectinate. Fore wing areolet absent; stigma relatively narrow (Fig. 1), Rs+2r arising near basal 0.3-0.4 of stigma. Hind wing (Fig. 4) with first abscissa of CU1 varying from equal in length to slightly shorter or longer than 1cu-a. T1 (Fig. 5) long, slender, parallel-sided from base to distinctly posteriorad spiracles, then abruptly expanding; weakly decurved in profile (Fig. 5); dorsal carinae extending full length of parallel-sided portion (to posterior end of S1), fading into and largely absent from expanded posterior portion; basal depression at dorsal tendon attachment small; dorsal-lateral carina complete between spiracle and apex of T1; glymma absent. S1 extending full length of parallel-sided portion (over basal 0.6, distinctly posteriorad spiracle). T2 thyridium absent (Fig 6); laterotergites of T2 and T3 separated by creases from median tergite. Ovipositor (Fig. 7) straight; ovipositor sheath more distinctly upcurved; ovipositor relatively shorter than in
Pion, abruptly becoming needle-like apically from distinctly swollen base (Fig. 7); without dorsal, subapical notch.
This description is based largely on S. minor and is expanded from that given for Syntactus by Townes (1970), who also included it in a key to genera of Pionini.