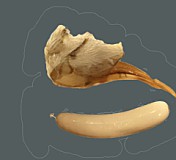
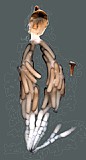
Clypeus highly variable among species and between sexes: varying from short and broad in some species to relatively tall and narrow in others; punctate, often coarsely so, especially apically, sometimes shagreened; ventral margin highly variable: sharp throughout in some species, thickened medially but sharp laterally in other species, when sharp, not distinctly impressed; shape of ventral margin either evenly convex, or broadly truncate medially and angled upwards laterally, or weakly concave in the middle of the truncate portion; also varying from sharp to blunt throughout, often blunt medially, sharp laterally; epistomal sulcus narrow, distinct, often sharply impressed, more rarely indistinct, sometimes varying between sexes; clypeus in profile flat to weakly bulging. Inner eye margins parallel. Malar space varying from slightly less than 0.5 times basal width of mandible to about 1.0 times basal width of mandible; malar sulcus absent. Mandible (Fig. 3) strongly curved, tapering from base to near middle then parallel-sided to weakly expanding from middle to apex; ventral tooth usually a little longer than dorsal tooth; ventral margin distinctly carinate. Maxillary palp shorter than height of head, less commonly equal to height of head; antenna (Figs 1, 3) usually slightly longer than body, first flagellomere moderate in length, typically as in Fig. 1 and 3. Ocelli small to moderate in size, diameter of lateral ocellus usually less than distance from lateral ocellus to eye. Hypostomal carina meeting occipital carina slightly but distinctly above base of mandible; occipital carina complete dorsally. Epomia sometimes absent or indistinct but present and distinct in at least three of the species examined. Epicnemial carina usually (95% of specimens examined) not reaching anterior margin of mesopleuron. Notaulus present usually as a weak impression on anterior declivity (the depression more rarely deeply impressed), weaker and indistinct on disk in some species, extending posteriorad level of tegula in other species though only rarely (one of the species examined) distinctly impressed. Groove between propodeum and metapleuron shallow, v-shaped, not u-shaped as in pionines; pleural carina present, usually strongly elevated; median longitudinal carinae very well-developed, forming flask-shaped median section with petiolar area broad and often delimited by median portion of posterior transverse carina; areola confluent with basal median area; lateral longitudinal carina weaker than median longitudinal carina, extending at least to spiracle from posterior margin and sometimes complete to anterior margin; posterior transverse carina complete in some species, completely absent in a few species; anterior transverse carina absent. Legs with apical margin of mid tibia either not expanded into a tooth or expanded into a tooth that is a little more rounded and not quite as well-developed as that of fore leg; apical comb on posterior side of hind tibia absent; posterior hind tibial spur 0.35-0.5 times length of long hind basitarsus (Figs 1-3); tarsal claws pectinate at extreme base in those species where the base was visible; fifth tarsomere of hing leg normal, not unusually elongate (relative to fourth) (Figs 1-3). Fore wing with areolet either present or absent, usually fairly consistent within a species; stigma moderately narrow as in Fig. 2, Rs+2r usually arising at or very slightly distad midpoint. Hind wing (Fig. 1) with first abscissa of CU1 either a little longer than, a little shorter than or equal in length to 1cu-a. T1 varying from relatively slender to somewhat broader: very gradually to more strongly expanding posteriorly; ventral margin usually weakly curved in profile; dorsal carinae absent or nearly so in the species examined; basal depression at dorsal tendon attachment varying absent or nearly so; dorsal-lateral carina varying from absent or nearly so in a few species to complete between spiracle and apex of T1 in some species; glymma absent. S1 varying among species: almost extending to level of spiracle in some species, extending to spiracle in others, and extending slightly posteriorad spiracle in some species. T2 thyridium not evident in species examined. Laterotergites of T2 and T3 separated by creases from median tergite. Ovipositor and sheath (Figs 1-3) downcurved or straight and posteriorly directed; ovipositor with distinct dorsal, subapical notch (Fig. 5); sheathes distinctive: outer surface of sheathes somewhat bulging, giving a slightly tubular appearance, usually broadest subbasally, truncate or nearly so at apex; hypopygium with distinctive pattern of short, erect setae. Male parameres elongate, extending distinctly beyond aedeagus in most individuals examined.
This description is based largely on about nine species in the Texas A&M University Collection and modified extensively from Townes (1970).