Head: Antenna filiform, longer than body. Frons, vertex, and temple smooth, shiny; frons bare, vertex and upper temple nearly so. Labrum exposed. Clypeus weakly to distinctly protruding in profile. Malar sulcus deeply impressed. Mandible gradually to somewhat more abruptly widening from apex to base, carinate ventrally over most of basal half, never with distinct basal tooth as in
Opius s. s. Maxillary palp longer than head, reaching mid coxa. Occipital carina present laterally, extending dorsal-medially at least to level of inner eye margin, broadly absent mid-dorsally; widely separated from hypostomal carina at base of mandible.
Mesosoma: Pronotum dorsally with narrow, transverse, crenulate sulcus extending continuously along lateral pronotum to ventral corner; weak to deep median pit interrupting sulcus dorsally. Mesoscutum in profile with anterior declivity very slightly concave, nearly vertical; notaulus present on anterior portion of mesoscutal disc, angled laterally at anterior end, laterally-directed portion carinate along anterior margin; notaulus continuous with weakly to distinctly crenulate impression bordering lateral mesoscutal margin, the impression extending posteriorly at least to level of tegula; midpit of mesoscutum well-developed, discrete. Scutellar sulcus narrow, though not exceptionally so, 3-4 times wider than mid-length, crenulate, with numerous closely-spaced ridges. Mesopleuron smooth, shiny, posterior margin not crenulate; precoxal sulcus distinctly impressed, crenulate. Propodeum with large median areola, variously obscured by sculpture.
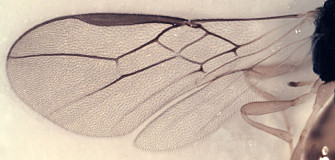
Wings: Slightly more than twice as long as wide. Stigma long, very narrow, nearly parallel-sided basally, widening distally; thickest part of stigma twice maximum width of proximal half. Radial cross-vein® thickened, weakly to strongly bowed anteriorly, arising from extreme base of stigma, nearly in line with 3RSa; RS+M weakly to distinctly sinuate; second submarginal cell nearly parallel-sided, not or only very weakly converging distally; m-cu nearly always postfurcal, entering second submarginal cell; 2CUa distinct, shorter than 2cu-a. Hind wing with both RS and M distinct nearly to wing margin, usually nebulous: very weakly pigmented; m-cu varying from indistinct in smaller individuals to present in larger individuals as a weakly pigmented impression extending more than half way to wing margin.
Legs: Hind femur slender, dorsal surface uneven, somewhat bilobed.
Metasoma: Petiole (T1) with distinct dorsope; dorsal and lateral surface densely striate to strigose, largely obscuring dorsal carinae except at base; spiracle of T1 located at or slightly posteriad mid-length; sternite short but distinct, extending 0.4-0.5 distance between base of T1 and spiracle. Metasoma posteriad petiole pyriform, unsculptured, with row of setae evenly spaced on posterior margin of tergites; T2 spiracle laterally displaced. Ovipositor nearly straight, without dorsal node; ovipositor sheath setose throughout.