Hypsantyx Pfankuch, 1906
Taxonomic History / Nomenclature
Hypsantyx Pfankuch, 1906: 92. Type species: Tryphon impressus Gravenhorst [= a junior subjective synonym of Hypstantyx lituratoria (Linnaeus, 1761)]. Monobasic.
Remarks
There is a single valid species, Hypsantyx lituratoria (Linnaeus, 1761). It occurs in Europe. Yu et al. (2012) give a long list of tenthredinid and diprionid hosts, plus associated references.
Diagnosis and Relationships

The only known species is robust and defined largely by the poorly developed epicnemial carina in combination with the blunt apical margin of the clypeus. Some of the Neotropical species of Phobetes also have a poorly developed epicnemial carina, but the fore wing is absent in these species and present in H. lituratoria. The mandibular teeth are also different in members of these two genera.
Description
Clypeus (Figs 2, 3) somewhat narrow, with surface finely punctate; ventral margin thickened, blunt and evenly convex throughout (Figs 2, 3); epistomal sulcus narrow, distinct; clypeus in profile essentially flat. Inner eye margins parallel. Malar space (Fig. 3) slightly more than 0.5 times basal width of mandible; malar sulcus absent. Mandible (Fig. 4) broad, gradually tapering from base to apex; dorsal tooth broader than ventral tooth, though both are broad and about equal in length; ventral margin distinctly carinate. Antenna (Fig. 5) equal to or slightly shorter than body, first flagellomere short relative to species in genera such as Mesoleptidea and Hadrodactylus, basal flagellomeres somewhat shorter and broader in female than male. Epomia not evident. Epicnemial carina greatly reduced, weak and only present ventrally when discernible, not reaching anterior margin of mesopleuron (Figs 7, 8). Notaulus (Fig. 6) present as a weakly impressed groove on anterior declivity. Propodeum (Fig. 9) with median longitudinal carinae very well-developed, forming flask-shaped median section with large rugulose petiolar area similar to that in Synomelix petiolar area and areola confluent but separated from basal median area by a distinct transverse carina; lateral longitudinal carina and transverse carinae weaker, the latter sometimes absent. Posterior hind tibial spur as in Fig. 1; fifth tarsomere of hing leg normal, not unusually elongate (relative to fourth). Fore wing (Fig. 6) with areolet present; stigma moderately broad, Rs+2r near basal third. Hind wing with first abscissa of CU1 approximately equal in length to 1cu-a. T1 (Figs 11-13) relatively short and broad, ventral margin nearly straight in profile; dorsal carinae distinctly elevated and extending posteriorly about half distance between spiracle and posterior margin; basal depression at dorsal tendon attachment broad, shallow; dorsal-lateral carina somewhat obscured by adjacent sculpture (Fig. 11); glymma absent. T2 thyridium readily visible (Fig. 14). Ovipositor and sheath (Fig. 14) straight, ovipositor with distinct dorsal, subapical notch.
The description is modified from Townes (1970), based on the images presented here from specimens in the Canadian National Collection.

1.
Hypsantyx lituratoria habitus male...
↴

2.
Hypsantyx lituratoria female fa...
↴

3.
Hypsantyx lituratoria face, m...
↴
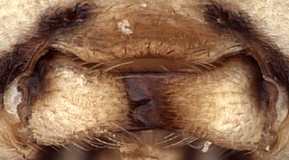
4.
Hypsantyx lituratoria mandibles...
↴

5.
Hypsantyx lituratoria antenna, female...
↴

6.Hypsantyx lituratoria

7.
Hypsantyx liturato...
↴
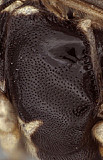
8.
Hypsantyx lit...
↴

9.
Hypsantyx lituratoria propode...
↴

10.
Hypsantyx lituratoria wings...
↴

11.
Hypsantyx lituratoria T1, femal...
↴

12.Hypsantyx lituratoria T1, female

13.
Hypsantyx lituratoria ...
↴

14.
Hypsantyx ...
↴

15.
Hypsantyx lituratoria ...
↴
Distribution
No referenced distribution records have been added to the database for this OTU.
Map
There are no specimens currently determined for this OTU, or those specimens determined for this OTU are not yet mappable.
Acknowledgements
This page was assembled by Bob Wharton as part of a larger collaborative effort on the genera of Ctenopelmatinae. Page last updated April, 2015.
This work would not have been possible without the groundwork provided by Ian Gauld’s study of the Australian and Costa Rican faunas, and we are particularly grateful for his assistance in many aspects of this study. We are especially thankful to Andy Bennett of the Canadian National Collection for extended loans of the material used for this webpage. We also thank David Wahl for useful feedback throughout our study. Matt Yoder provided considerable assistance with databasing issues, and our use of PURLs (http://purl.oclc.org) in this regard follows the example of their use in publications by Norm Johnson. Heather Cummins, Caitlin Nessner, Karl Roeder, Danielle Restuccia, and Cheryl Hyde graciously assisted us with image processing, formatting, and literature retrieval. This study was supported by the National Science Foundation’s PEET program under Grant No. DEB 0328922 and associated REU supplement nos DEB 0723663 and 0923134.
This material is based upon work at Texas A&M University supported by the National Science Foundation under Grant Number DEB 0328922 with REU supplements DEB 0723663 and 0923134… Any opinions, findings, and conclusions or recommendations expressed in this material are those of the author(s) and do not necessarily reflect the views of the National Science Foundation.
