Gastroporus Foerster, 1869: 206 Type species: Hyperallus caliroae Viereck, 1911. Subsequent inclusion by Perkins (1962: 424). Synonymized by Perkins (1962: 424).
Hyperallus Foerster, 1868
Taxonomic History / Nomenclature
Hyperallus Foerster, 1869: 210 Type species: Hyperallus caliroae Viereck, 1911: 189-190. Subsequent inclusion by Viereck (1911: 189). Monobasic.
Remarks
Until recently, Hyperallus was known only from the type species from Eastern United States. Kasparyan (in Kasparyan and Khalaim 2007) relatively recently described a second species from the Eastern Palaearctic region. Specimens in the Texas A&M University collection from Ontario (Canada) and Michigan and Texas (USA) represent new but not surprising distribution records for North America. The type species is variable in color, as indicated by the Figs 2-4 vs. 19-22. There is not sufficient material at hand to determine if the color variation is limited to sexual dimorphism. The two specimens from Texas are larger and have more extensive orange coloration than the two primary type specimens and the non-type material from Michigan and Canada.
Diagnosis and Relationships

The species of Hyperallus are recognized by the following combination of characters: blunt, evenly convex margin of the clypeus, absence of a fore wing areolet, and well-developed propodeal carinae with large, well-delimited petiolar area. The species of Synomelix have a similar arrangement of propodeal carinae, but the apex of the clypeus is sharp. The species of Phobetes also have a blunt clypeus but the propodeal carinae are poorly developed. The species of Hypamblys are most similar to those of Hyperallus but in Hypamblys T1 tends to be a little longer and narrower, the extreme lateral margin of the clypeus is usually sharp or impressed, and the propodeal carinae, though more developed than in Phobetes, tend to be less developed than in Hyperallus.
Description
Clypeus (Figs 5, 6, 19) moderate in width, with surface finely punctate; ventral margin blunt and evenly convex throughout; epistomal sulcus narrow, somewhat indistinct; clypeus in profile flat. Inner eye margins parallel. Malar space (Figs 5, 19) short, equal to or slightly less than 0.5 times basal width of mandible; malar sulcus absent. Mandible (Figs 6, 19) tapering from base to apex; ventral tooth a little longer than dorsal tooth; ventral margin distinctly carinate. Maxillary palp shorter than height of head; antenna (Figs 1, 7, 19, 20) equal to or slightly shorter than body, first flagellomere (Fig. 7) short relative to species in most other genera of Euryproctini. Ocelli small to moderate in size, diameter of lateral ocellus a little less than distance from lateral ocellus to eye. Hypostomal carina meeting occipital carina distinctly above base of mandible; occipital carina complete dorsally. Epomia absent. Epicnemial carina ( Fig. 8) not reaching anterior margin of mesopleuron. Notaulus (Figs 9-11) present usually as a deep, distinct impression on anterior declivity, becoming distinctly weaker and shallow on disk, extending to or just posteriorad level of tegula. Groove between propodeum and metapleuron absent to very weakly indicated, not u-shaped as in pionines; pleural carina present, strongly elevated; propodeum with posterior fields distinctly declivous (Fig. 12); median longitudinal carinae very well-developed, forming flask-shaped median section with petiolar area (Fig. 13) broad and distinctly separated from small areola by a transverse carina; lateral longitudinal carina usually distinct between posterior transverse carina and spiracle; posterior transverse carinae complete and well-developed. Legs (Figs 1, 2) with apical comb on posterior side of hind tibia present though not strongly developed; posterior hind tibial spur about 0.5 times length of hind basitarsus; tarsal claws not pectinate; fifth tarsomere of hing leg normal, not unusually elongate (relative to fourth). Fore wing (Figs 2, 14, 21) with areolet absent; stigma very broad, Rs+2r usually arising from midpoint. Hind wing (Fig. 14) with first abscissa of CU1 longer than 1cu-a. T1 (Figs 4, 15, 16, 20) broad; ventral margin straight in profile; dorsal carinae present, varying from low, barely extending to level of spiracles to more distinctly elevated and extending posteriorad spiracles; basal depression at dorsal tendon attachment varying broad, very shallow; dorsal-lateral carina complete between spiracle and apex of T1; glymma absent. S1 not extending to level of spiracle, only about 0.35 times length of T1. T2 thyridium present. Laterotergites of T2 and T3 separated by creases from median tergite. Ovipositor and sheath (Figs 1, 17, 18) straight, ovipositor with deep, broad dorsal, subapical notch.
The above description is considerably modified from Townes (1970), and based on the holotypes of Polyterus caliroae Rohwer and Hyperallus caliroae Viereck as well as four specimens in the Texas A&M University collection.

1.
Hyperallus habitus ...
↴

2.
Holotype Hyperallus cali...
↴

3.
Holotype Hyperallus cali...
↴

4.
Holotype Hyperallus cali...
↴
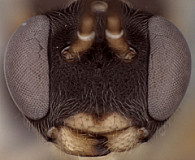
5.Hyperallus face

6.Hyperallus mandibles

7.
Hyper...
↴

8.Hyperallus mesosoma lateral
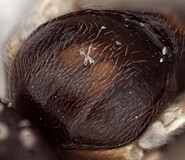
9.
Hyperallus caliroae mesoscut...
↴
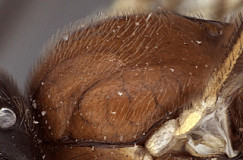
10.Hyperallus mesoscutum

11.
Hyperallus caliroae mesoscutum...
↴

12.
Hyperallus propode...
↴

13.Hyperallus propodeum

14.Hyperallus wings

15.
Hyperallus caliroa...
↴

16.Hyperallus T1

17.Hyperallus ovipositor
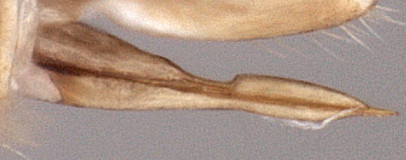
18.Hyperallus ovipositor

19.
Holotype Polyterus calir...
↴

20.
Holotype Polyterus calir...
↴

21.
Holotype Polyterus calir...
↴

22.
Holotype Polyterus calir...
↴
Distribution
No referenced distribution records have been added to the database for this OTU.
Biology / Hosts
The holotype of Hyperallus caliroae was reared from the sawfly Caliroa liturata MacGillivray (Tenthredinidae).
Map
There are no specimens currently determined for this OTU, or those specimens determined for this OTU are not yet mappable.
Acknowledgements
This page was assembled by Bob Wharton as part of a larger collaborative effort on the genera of Ctenopelmatinae. Page last updated April, 2015.
This work would not have been possible without the groundwork provided by Ian Gauld’s study of the Australian and Costa Rican faunas, and we are particularly grateful for his assistance in many aspects of this study. We thank David Wahl of the American Entomological Institute, Gavin Broad of The Natural History Museum, London, Bob Kula of the USDA Systematic Entomology Laboratory, and Andy Bennett of the Canadian National Collection for extended loans of the material used for this study. We also thank David Wahl for useful feedback throughout our study. Matt Yoder provided considerable assistance with databasing issues, and our use of PURLs (http://purl.oclc.org) in this regard follows the example of their use in publications by Norm Johnson. Heather Cummins, Andrea Walker, Patricia Mullins, Caitlin Nessner, Mika Cameron, Karl Roeder, Danielle Restuccia, and Cheryl Hyde graciously assisted us with image processing, formatting, and literature retrieval. This study was supported by the National Science Foundation’s PEET program under Grant No. DEB 0328922 and associated REU supplement nos DEB 0616851, 0723663, 0822676, 0923134, and 1026618.
This material is based upon work at Texas A&M University supported by the National Science Foundation under Grant Number DEB 0328922 with REU supplements DEB 0616851, 0723663, 0822676, 0923134, and 1026618.. Any opinions, findings, and conclusions or recommendations expressed in this material are those of the author(s) and do not necessarily reflect the views of the National Science Foundation.
