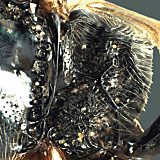
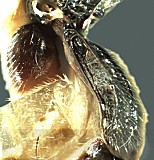
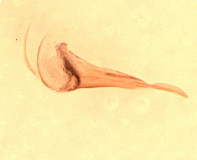
Hodostates is presently included in the tribe Pionini. It is distinguished from all other Pionini by the combination of 1) distinct subapical notch on the ovipositor (Figs 14, 16); 2) completely areolate propodeum (Fig. 11) with posterior field sharply declivitous (Figs 9, 10); 3) presence of a distinct glymma subbasally (Fig. 12); and 4) long notauli extending posteriorly as distinctly impressed grooves at least over anterior 0.75 of mesoscutum (Fig. 8).
Hodostates also resembles certain euryproctine genera with a short, areolate propodeum and a broader T1, but differs from all euryproctines in the possession of a glymma.
Hodostates differs from mesoleiine genera by the combination of a completely areolate propodeum, evenly margined clypeus (Figs 5, 6), presence of fore wing areolet (Figs 3, 7), and the exceptionally long, distinctly impressed notauli (Fig. 8).
Hodostates can be distinguished from all Perilissini by the combination of the small glymma, deep notauli and clypeus without distinctly impressed margin.
Figures 1 and 9 are of the lectotype of H. brevis; Figs 2, 4, 6, 7, and 10 are of the holotype of palustris Habermehl, a synonym of brevis; Figs 3, 11, and 14 are of the lectotype of H. rotundatus; Figs. 5, 12, and 13 are of other determined specimens of H. rotundatus; Fig. 8 is of the holotype of caliroae Rohwer, a synonym of rotundatus; Figs 15 and 16 are of another determined specimen of H. brevis.
1.
Hodostates brevis Thomso...
Hodostates brevis Thomson lateral habitus
↰
↴
2.
Hodostates brevis dorsal...
Hodostates brevis dorsal view.
↰
↴
3.
Hodostates rotundatus (Davis) ...
Hodostates rotundatus (Davis) lateral habitus.
↰
↴
4.
Hodostates brevis dor...
Hodostates brevis dorsal view of head.
↰
↴
5.
Hodostates rotundatus ...
Hodostates rotundatus face.
↰
↴
6.
Hodostates brevis clypeus and ...
Hodostates brevis clypeus and mandibles.
↰
↴
7.Hodostates brevis wings.
8.
Hodostates rotundatus sh...
Hodostates rotundatus showing elongate notauli.
↰
↴
9.
Hodostates brevis showin...
Hodostates brevis showing sharply declivitous posterior field of propodeum.
↰
↴
10.
Hodostates brevis propod...
Hodostates brevis propodeum lateral view.
↰
↴
11.
Hodostates rotundatus pr...
Hodostates rotundatus propodeum and first metasomal tergite, dorsal view.
↰
↴
12.
Hodostates rotundatus ...
Hodostates rotundatus first tergite with glymma.
↰
↴
13.
Hodostates rotundatus ov...
Hodostates rotundatus ovipositor sheath showing color variation of metasoma.
↰
↴
14.
Hodostates rotundatus oviposito...
Hodostates rotundatus ovipositor.
↰
↴
15.
Hodostates brevis ventra...
Hodostates brevis ventral view showing hypopygium and barely protruding ovipositor sheath.
↰
↴
16.
Hodostates brevis dorsal valve...
Hodostates brevis dorsal valve of ovipositor showing subapical notch.
↰
↴