Anoncus Townes, 1970
Taxonomic History / Nomenclature
Anoncus Townes, 1970: 127. Type species: Mesoleius striatus Davis, 1898. Original Designation.
Remarks
21 valid species are listed by Yu et al. (2012). Townes, 1970: 127 states there are about 50 species in the American Entomological Institute Collections, most of which are undetermined or undescribed. Apparently limited to the Holarctic Region.
Diagnosis and Relationships

Anoncus is apparently unique among the Mesoleiini in the possession of an ovipositor without a subapical, dorsal notch. This condition is found in other tribes of Ctenopelmatinae, notably Pionini, a few Perilissini, and several Ctenopelmatini. T1 and T2 in Anoncus are typical of Mesoleiini and Anoncus also lacks the broad u-shaped groove between the metathorax and the lateral longitudinal carina of the propodeum that characterizes most pionines.
Description
Frons without median horn or elevated carina. Clypeus (Figs 3, 4) short and wide, strongly bulging medially and subapically; ventral margin sharp except medially where sharp margin overlapped by blunt median bulge (Fig. 4); ventral margin usually truncate, with sharp lateral margins distinctly angled dorsally; epistomal sulcus sharp. Malar space shorter than half basal width of mandible (Fig. 3), very short in some individuals. Mandible (Fig. 4) moderately long, curved, gradually narrowing from base to apex or more or less parallel-sided over apical half, ventral tooth varying from about equal in length to distinctly longer (Fig. 4) than dorsal tooth, when equal, dorsal tooth often slight broader. Inner eye margins parallel (Fig. 3) to weakly converging. Ocelli small, with maximum diameter of lateral ocellus distinctly shorter than distance between ocellus and eye. Female and male antennae longer than body; first flagellomere long and slender, nearly twice longer than second (Figs 1, 5). Hypostomal carina joining occipital carina well above base of mandible; occipital carina complete. Epomia absent or indistinct due to heavy sculpture (Fig. 6). Dorsal end of epicnemial carina nearly always extending to anterior margin of mesopleuron (Fig. 7); mesopleuron mat ventrally, but only faintly so in some species and more rugulose in others. Notaulus (Figs 8, 9) sharply impressed on anterior declivity (though not quite reaching anterior margin), weak to indistinct on disk. Pleural carina complete, usually well-developed; lateral longitudinal carina of propodeum nearly always well-developed, complete or nearly so; median longitudinal carinae forming a broad, rounded petiolar area then extending anteriorly as very narrowly spaced ridges or confluent and forming a weaker, irregular median ridge; short, broad petiolar area separated by transverse carina from narrow areola confluent with basal median area; otherwise, anterior and posterior transverse carinae absent. Legs usually with apical comb on posterior side of hind tibia poorly developed; hind tibial spurs long, slender (Figs 2, 11); all tarsal claws simple, not pectinate. Fore wing (Fig. 12) with areolet absent; stigma not exceptionally broad, with Rs+2r arising from midpoint. Hind wing (Fig 12) with first abscissa of CU1 much longer than 1cu-a. T1 (Figs 13, 14) not long and slender; gradually widening posteriorly; basal depression for dorsal tendon attachment deep; dorsal carinae usually well-developed, extending from base to level of spiracle or somewhat more posteriorly, but weakening posteriorly; dorsal-lateral carina usually sharp and distinct from spiracle to apex; glymma deep, broad basally, narrowing posteriorly. S1 extending to level of spiracle or nearly so. T2 thyridium present; laterotergites of T2 and T3 completely separated by creases, though T3 crease not well developed posteriorly in the material examined. Ovipositor fairly short (Figs 1, 15, 16), straight, completely lacking subapical, dorsal notch (Figs 15, 16) typical of all other mesoleiines, though slightly swollen as a minute node in some species; ovipositor sheath narrow. Apex of female metasoma as in Fig. 15.
According to Townes, 1970: 127," segments 2-4 of middle and hind tarsi are exceptional in tending to be short, enlarged apically, and with conspicuous apical bristles (especially in females)" However, this is set of features is also found in other mesoleiines and may not be as exceptional as indicated.
This description is modified from that of Townes, 1970: 127 and based on three species in the Texas A&M University Collection.

1.
Anoncus referendus habitus...
↴

2.Anoncus habitus
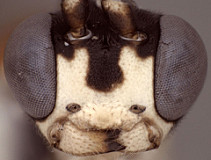
3.Anoncus face and clypeus

4.
Anoncus mandibles and clypeus...
↴

5.
Anoncus antenn...
↴
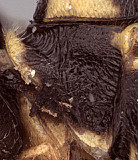
6.
Anoncus pronotum...
↴

7.
Anoncus mesopleuro...
↴

8.
Anoncus mesoscutum...
↴

9.
Anoncus mesoscutum...
↴

10.Anoncus propodeum

11.Anoncus hind leg

12.Anoncus wings

13.
Anoncus T1...
↴

14.

15.
Anoncus ovipositor and s...
↴

16.
Anoncus ovipositor...
↴
Distribution
No referenced distribution records have been added to the database for this OTU.
Biology / Hosts
Recorded hosts are tenthredinid sawflies (Yu et al. 2012), and at least one species attacks gall-forming sawflies on willows (Salix). Kasparyan and Kopelke (2009) reviewed ichneumonid parasitoids of gall-forming sawflies on willows.
Map
There are no specimens currently determined for this OTU, or those specimens determined for this OTU are not yet mappable.
Acknowledgements
This page was assembled by Bob Wharton as part of a larger collaborative effort on the genera of Ctenopelmatinae. Page last updated May, 2015.
This work would not have been possible without the groundwork provided by Ian Gauld’s study of the Australian and Costa Rican faunas, and we are particularly grateful for his assistance in many aspects of this study. We also thank David Wahl of the American Entomological Institute, Andy Bennett of the Canadian National Collection, and Gavin Broad of The Natural History Museum, London for extended loans of the material used for this study and Dave Karlsson for access to material from the Swedish Malaise Trap Survey (trap 10, collection event 398). Matt Yoder provided considerable assistance with databasing issues, and our use of PURLs (http://purl.oclc.org) in this regard follows the example of their use in publications by Norm Johnson. Heather Cummins, Andrea Walker, Patricia Mullins, Caitlin Nessner, Mika Cameron, Karl Roeder, Amanda Ladigo, and Cheryl Hyde graciously assisted with image processing, formatting, and literature retrieval. This study was supported by the National Science Foundation’s PEET program under Grant No. DEB 0328922 and associated REU supplement #s DEB 0723663, 0923134, and 1026618.
This material is based upon work at Texas A&M University supported by the National Science Foundation under Grant Number DEB 0328922 with REU supplements DEB 0723663, 0923134, and 1026618. Any opinions, findings, and conclusions or recommendations expressed in this material are those of the author(s) and do not necessarily reflect the views of the National Science Foundation.
