Nanium Townes, 1967
Taxonomic History / Nomenclature
Nanium Townes, 1967: 174-175. Type species: Tryphon capitatus Cresson (1864). Original designation.
Remarks
The genus Laphyroscopus was proposed by Foerster (1869) without included species. Davis (1897) was the first to include a species in Laphyroscopus, placing Tryphon capitatus Cresson there. Viereck (1914) recognized the placement by Davis as the designation of a type species (monobasic) for Laphyroscopus. However, current workers generally accept that many of the names proposed by Thomson are unjustified emendations of the Foerster names. Consequently, Tryphon gorskii Ratzeburg, is recognized as the type species of Laphyroscopus based on the placement of gorskii in Luphyroscopus by Thomson (1883).
There are five valid species as of March, 2011:
Nanium capitatum (Cresson, 1864)
Nanium huberthi Gauld,1997
Nanium mairenai Gauld,1997
Nanium nogueri Gauld,1997
Nanium oriasi Gauld,1997
Diagnosis and Relationships

The species of Nanium are generally small-bodied, similar in size to the smaller species of Lathrolestes. Defining features (see Fig. 1) include the absence of propodeal carinae (except for occasional traces of median longitudinal carinae posteriorly) and granulate sculpture on the propodeum, T1, and (in most but not all species) the mesoscutum. Also, unlike many of the perilissine genera, the epicnemial carina extends to the anterior margin of the mesopleuron and the stigma is short and very deep. The marginal cell is also short and deep. The mediella tends to be more sharply bulging than most perilissines, but this character may be size-related.
There is a fair amount of diversity among the undescribed neotropical species that have been accumulating in various museums. A revision of the neotropical genera of Perilissini is badly needed. The above description is based on numerous specimens of Nanium capitatum from various localities in the United States plus several undetermined specimens from Mexico and at least three different species from South America.

1.
Nanium showing sculpture...
↴
Description
Clypeus (Figs 3, 4) with ventral margin thick and bluntly rounded, thinner laterally, without small lateral tooth or projection; ventral margin convex; epistomal sulcus present, usually distinct (Figs 3, 4), clypeus in profile not projecting or only very weakly angled outwardly from face. Malar space distinct, very short (0.2 x basal width of mandible) to moderately long (0.3-0.5 x basal width of mandible). Mandible (Fig. 4) with ventral tooth equal to or very slightly longer than dorsal tooth. Lateral ocellus much shorter than distance between ocellus and eye. Maxillary palp moderately long, about equal to head height; female antennae varying from as long as to slightly but distinctly longer than body; first flagellomere with discrete tyloid containing fewer than 15 sensilla. Hypostomal carina joining occipital carina well before base of mandible; occipital carina complete. Dorsal end of epicnemial carina extending to anterior margin of mesopleuron. Notaulus varying from nearly absent to distinctly impressed on anterior declivity, extending posteriorly to level of tegula as a weak groove. Groove between propodeum and metanotum in lateral view indistinct; pleural carina complete, well-developed in type species, absent or nearly so in some undescribed neotropical species; propodeal carinae completely absent (Fig. 6). Apex of mid tibia somewhat evenly rounded, with at most a small, weak tooth; tooth at apex of fore tibia also often weakly developed; apex of hind tibia with poorly developed comb: setae widely spaced, not dense; posterior hind tibial spur at least 7x longer than maximum width at base; tarsal claws varying from completely pectinate to pectinate only on basal half. Fore wing areolet nearly always absent; stigma short, broad, Rs+2r arising near middle of stigma. Hind wing with distal abscissa of CU1 not always discernible but when visible, first abscissa of Cu1 distinctly longer than 1cu-a. T1 varying from moderately long and slender to shorter, more distinctly broadening posteriorly; flattened dorsally, without discrete dorsal carinae; without basal depression at dorsal tendon attachment; dorsal-lateral carina absent or nearly so between spiracle and apex of T1; glymmae on each side meeting on the midline posterior to dorsal tendon attachment, large, deep, separated at midline by translucent partition. T2 thyridium absent; laterotergites of T2 and T3 not or only partially separated by creases in most specimens examined. Ovipositor straight, short, with very broad, deep, subapical depression (Fig. 7); ovipositor sheath narrowly rectangular to weakly expanding distally, bluntly rounded apically. Male parameres broad, somewhat rectangular, never strongly narrowed posteriorly; aedeagus rounded and weakly clubbed distally.


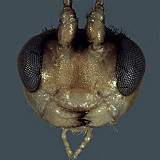
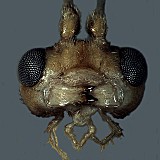


Distribution
This is an New World genus, found in the eastern Nearctic region from southern Canada to Texas and throughtout the Neotropical region as far south as Chile and Argentina.
Distribution
No referenced distribution records have been added to the database for this OTU.
Biology / Hosts
Hosts and biology unknown. Gauld (1997) suggested that hosts might be one of the smaller pergid sawflied.
Map
There are no specimens currently determined for this OTU, or those specimens determined for this OTU are not yet mappable.
Acknowledgements
This page was assembled by Bob Wharton as part of a larger collaborative effort on the genera of Ctenopelmatinae. This work would not have been possible without the groundwork provided by Ian Gauld’s study of the Australian and Costa Rican faunas, and we are particularly grateful for his assistance in many aspects of this study. We also thank David Wahl for useful feedback throughout our study and to Gavin Broad for exchange of information on Perilissini. Matt Yoder provided considerable assistance with databasing issues, and our use of PURLs (http://purl.oclc.org) in this regard follows the example of their use in publications by Norm Johnson. Patricia Mullins, Amy James, Heather Cummins, and Caitlin Nessner graciously assisted with image capture, processing, and formatting. This study was supported by the National Science Foundation’s PEET program under Grant No. DEB 0328922 and associated REU supplement No. DEB 0723663 and RET supplement Nos. 0723663 and 0822676. Page last updated March, 2011.
This material is based upon work supported by the National Science Foundation under Grant Number DEB 0328922 with REU supplement DEB 0723663 and RET supplements number 0723663 and 0822676.
Any opinions, findings, and conclusions or recommendations expressed in this material are those of the author(s) and do not necessarily reflect the views of the National Science Foundation.
