Saotus Thomson, 1883: 933 Unjustified emendation.
Iskarus Kolarov, 1987. Type species: Iskarus seleuciformis Kolarov, 1987. Monobasic. Synonymized by Kasparyan and Shaw (2003).
Saotus Thomson, 1883: 933 Unjustified emendation.
Iskarus Kolarov, 1987. Type species: Iskarus seleuciformis Kolarov, 1987. Monobasic. Synonymized by Kasparyan and Shaw (2003).
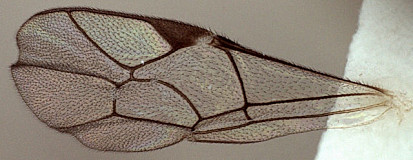
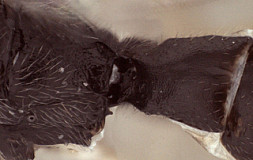
This description is considerably modified from Townes (1970) and based largely on females representing five species in the Texas A&M University Collection.
There are no specimens currently determined for this OTU, or those specimens determined for this OTU are not yet mappable.
This work would not have been possible without the groundwork provided by Ian Gauld’s study of the Australian and Costa Rican faunas, and we are particularly grateful for his assistance in many aspects of this study. We are also grateful to David Wahl of the American Entomological Institute and Andy Bennett of the Canadian National Collection for extended loans of the material used for this study. We also thank David Wahl for useful feedback throughout our study as well as Al Gillogly for collecting several species useful for composing this page and Dave Karlsson for access to material from the Swedish Malaise Trap Survey (Trap 22, collection event 1638 for S. compressiuscula). Matt Yoder provided considerable assistance with databasing issues, and our use of PURLs (http://purl.oclc.org) in this regard follows the example of their use in publications by Norm Johnson. Heather Cummins, Caitlin Nessner, Mika Cameron, Karl Roeder, Danielle Restuccia, and Cheryl Hyde graciously assisted us with image processing, formatting, and literature retrieval. This study was supported by the National Science Foundation’s PEET program under Grant No. DEB 0328922 and associated REU supplement nos DEB 0723663 and 0923134.
This material is based upon work at Texas A&M University supported by the National Science Foundation under Grant Number DEB 0328922 with REU supplements DEB 0723663 and 0923134. Any opinions, findings, and conclusions or recommendations expressed in this material are those of the author(s) and do not necessarily reflect the views of the National Science Foundation.
The keys, text, and images presented here were initially developed as part of an NSF-sponsored PEET project, DEB0328922. This page is a joint project of NSF PEET grants NSF DEB-0328922; ideas and opinions expressed here are those of the authors, and not the NSF; best viewed with Firefox, Mozilla, or OS X Safari. This material is copyright Bob Wharton (2007 and onwards) and may not be reproduced without permission.