Synonym: Parachasma Fischer, 1967 (Fischer 1977).
In many of the publications prior to 1980, the species of Doryctobracon were placed either in the genus Opius or in the genus Biosteres. The name Parachasma was also used for about 10 years prior to the discovery by Fischer (1977) that Doryctobracon, previously included in a different subfamily, was the same as Parachasma.Doryctobracon Enderlein, 1920
Taxonomic History / Nomenclature
Doryctobracon Enderlein, 1920: 144. Type species: Doryctobracon conjungens Enderlein, 1920 (original designation).
Description

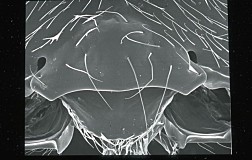
Occipital carina absent (Figs. 1, 2). Labrum varying from almost completely concealed to partially but distinctly exposed below ventral margin of clypeus; ventral margin of clypeus varying from very weakly sinuate, nearly truncate (Fig. 7), to strongly sinuate and produced ventrally to form a distinct median tooth or lobe. First flagellomere almost always shorter than second, and first flagellomere with dense patch of close-set, curved placode sensilla on outer side. Propleuron always without oblique carina dorsad propleural flange. Notauli deep at anterior margin in nearly all species, weak to absent posteriorly, rarely meeting in a weak, median pit (midpit usually absent); notauli always unsculptured. Postpectal carina absent. Hind tibia dorso-posteriorly without basal carina. Fore wing (Fig. 3) with second submarginal cell short; m-cu arising basad or directly in line with 2RS, only very rarely (1% of specimens in certain species) arising distad 2RS. Hind wing (Fig. 4) with RS absent basally, represented only by an infumate crease distally in a few species; m-cu long, well-developed though not tubular, nearly reaching wing margin, usually represented by a well-pigmented crease. Metasomal terga always unsculptured beyond petiole. Ovipositor always very long (Figs. 5, 6). For additional details, see Wharton (1997).






Diagnosis and Relationships
Doryctobracon is readily distinguished from other Opiinae with small second submarginal cells by the absence of an occipital carina, the sinuate margin of the clypeus, relatively reduced notauli, and the origin of fore wing m-cu basad or directly in line with 2RS. The species of Doryctobracon also have shorter, thicker flagellomeres clothed with shorter, thicker setae than do nearly all species of Diachasmimorpha, Fopius, Rhynchosteres, and Psyttalia. Further, they have a shelf formed behind a carinate flange where the anterior declivity of the mesonotum meets the pronotal collar dorsally.
Unless you are already on this page, considerably more information on species groups and use in biological control can be found on the Doryctobracon page (and associated links) of the Paroffit website page: http://mx.speciesfile.org/projects/8/public/public_content/show/13335?content_template_id=88
Biology / Hosts
The best known and most widespread species, D. crawfordi (Viereck) and D. areolatus (Szépligeti), attack tephritid pests (primarily members of the genus Anastrepha) in a variety of cultivated and native edible fruits in Latin America. There are also several other species that are known to attack fruit-infesting tephritids (Wharton and Marsh 1978). However, at least one species attacks tephritids in stems and one other species, Doryctobracon anneae Wharton, is only known to attack a species of tephritid breeding in flower heads of Dahlia spp. (Asteraceae) (Wharton and Norrbom 2013).
Biology and Behavior
Available information suggests that all species of Doryctobracon attack late instar larvae of their host tephritids; and as in all other Opiinae, they emerge from the host puparium.
Males of the species of Doryctobracon examined thus far share with Fopius and Diachasmimorpha paired Hagen’s glands with exceptionally long, thin filaments. These glands occupy much of the lateral portions of the abdominal cavity. For further comments about these glands, see Diachasmimorpha page.
Distribution
Doryctobracon is known only from the New World, ranging from extreme southern United States (Florida and a few old records from Texas) through Mexico and much of the Caribbean to northern Argentina. There are no records from Chile. As of 2012, there are also no records of successful biological control introductions outside the New World, though there have been some attempts. Thus, at present, Doryctobracon is confined to the Nearctic and Neotropical regions. See Ovruski et al. (2000) for summary of published information.
Distribution
No referenced distribution records have been added to the database for this OTU.
Map
There are no specimens currently determined for this OTU, or those specimens determined for this OTU are not yet mappable.
Acknowledgements
This page was assembled by Bob Wharton and Danielle Restuccia. It is part of a review of the genera of World Opiinae, conducted at Texas A&M University. We are particularly grateful to Xanthe Shirley, Andrew Ly, Patricia Mullins, Trent Hawkins, Lauren Ward, Cheryl Hyde, Karl Roeder, and Andrea Walker, who did nearly all of the imaging (together with Danielle) for this project. Matt Yoder and Istvan Miko provided guidance on databasing issues associated with our use of mx and HAO respectively. This project would not have been possible without the kindness of many curators at museums throughout the world who gave generously of their time to Bob Wharton and his students. In particular, I thank Henry Townes (deceased) and David Wahl (American Entomological Institute, Gainesville), Gordon Nishida (Bernice P. Bishop Museum, Honolulu), Norm Penny, and Bob Zuparko (California Academy of Sciences, San Francisco), Bill Mason (deceased), Mike Sharkey, Andrew Bennett, and Henri Goulet (Canadian National Collection, Ottawa), Paul Dessart (deceased) (Institut Royal des Sciences Naturelles de Belgique, Brussels), Marc De Meyer (Koninklijk Museum voor Midden-Afrika, Tervuren), Axel Bachmann (Museo Argentino de Ciencias Natureles, Buenos Aires), Eberhard Koenigsmann (deceased) and Frank Koch (Museum fuer Naturkunde der Humboldt-Universitaet, Berlin), J. Casevitz Weulersse and Claire Villemant (Museum National d’Historie Naturelle, Paris), James O’Connor (National Museum of Ireland, Dublin), Jenö Papp (National Museum of Natural History, Budapest), Kees van Achterberg (National Museum of Natural History, Leiden), Max Fischer, Herb Zettel, and Dominique Zimmermann (Naturhistorisches Museum, Wien), Per Persson and Lars-Åke Janzon (Naturhistoriska Riksmuseet, Stockholm), Ermenegildo Tremblay (Silvestri Collection, Portici), Erasmus Haeselbarth (Staatliche Naturwissenschaftliche Sammlungen Bayerns, Munich), Tom Huddleston and Gavin Broad (The Natural History Museum, London), Paul Marsh and Robert Kula (USDA Systematic Research Laboratory and US National Museum of Natural History, Washington, D. C.), Vladimir Tobias (deceased) and Sergey Belokobylskij (Zoological Institute, Academy of Sciences, St. Petersburg), and Roy Danielsson (Zoological Institute, Department of Systematics, Lund) for facilitating loans and general assistance associated with examination of holotypes and other material in their care. This work was supported largely by NSF/PEET DEB 0328922 and 0949027, with REU supplements 0723663, 1026618, 1213790, and 1313933 (to Wharton). Page last updated July, 2015. The material on this page is freely available, but should be acknowledged if used elsewhere.
This material is based upon work supported by the National Science Foundation under Grant Numbers DEB 9300517, DEB (PEET) 9712543, DEB (PEET) 0328922 with REU supplements 0723663 and 1026618 and DEB 0949027 with REU supplements 1213790 and 1313933. Any opinions, findings, and conclusions or recommendations expressed in this material are those of the author(s) and do not necessarily reflect the views of the National Science Foundation.
