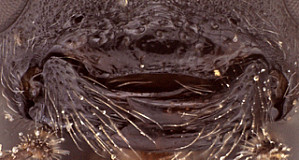
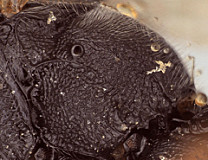
Clypeus (Fig. 2) short and relatively wide, weakly bulging subapically (nearly flat), with rounded transverse ridge; ventral margin sharp throughout, slightly impressed medially and distinctly visible throughout; ventral margin truncate medially, with lateral margins distinctly angled dorsally; epistomal sulcus distinct. Malar space 0.5-1.0 times basal width of mandible in female, shorter than half basal width of mandible in male. Mandible (Fig. 2) long, curved, gradually narrowing from base to apex; ventral tooth distinctly longer than dorsal tooth. Inner eye margins parallel. Ocelli small, with maximum diameter of lateral ocellus a little shorter than distance between ocellus and eye. Female (Fig. 1) and male antennae as long as or slightly longer than body; first flagellomere long and slender. Hypostomal carina joining occipital carina well above base of mandible; occipital carina complete. Epomia absent. Dorsal end of epicnemial carina weakening as it approaches anterior margin of mesopleuron, usually not extending to anterior margin (Fig. 3); entire mesopleuron except for speculum coarsely sculptured (Fig. 3): wrinkled to rugulose-mat. Notaulus absent to short and weakly impressed. Pleural carina (Fig. 4) usually distinct dorsoanteriorly, weak to absent ventroposteriorly; propodeal carinae usually weak: lateral longitudinal carina often poorly developed (Fig. 4); petiolar area completely to largely delimited by carinae; median longitudinal carinae extending irregularly and narrowly spaced from base of petiolar area to anterior margin, sometimes obscured by ruguose background sculpture of propodeum. Legs with apical comb on posterior side of hind tibia well developed, dense, though sometimes short; hind tibial spurs long, slender (Fig. 1), longest spur less than half length of long hind basitarsus in the material examined; all tarsal claws apparently simple, not pectinate. Fore wing (Fig. 5) with areolet usually present, usually small, sometimes with one side replaced by an unpigmented vein or bulla; stigma not exceptionally broad, with Rs+2r arising a little basad midpoint. Hind wing (Fig. 5) with first abscissa of CU1 longer than 1cu-a. T1 (Figs 6-7) relatively long and narrow, gradually widening posteriorly; dorsal carinae usually extending as narrowly spaced ridges posteriorad level of spiracle, often reaching about half distance between spiracle and posterior margin; basal depression of dorsal tendon attachment deep, wide; dorsal-lateral carina sharp and distinct from spiracle to posterior margin of T1, often poorly indicated in immediate vicinity of spiracle; glymma deep, broad basally, narrowing posteriorly. S1 not extending to level of spiracle. T2 thyridium present; laterotergites of T2 and T3 completely separated by creases. Ovipositor short, with subapical, dorsal notch.
This description is modified from Townes (1970) and based largely on one species in the Texas A&M University Collection.